

On this page:
Introduction The Two Transport Systems in Plants Transpiration The Transpiration Stream Structure of Xylem Factors Affecting Transpiration Rate Adaptations to Reduce Water Loss Translocation Sources and Sinks The Pressure Flow Hypothesis Structure of Phloem Comparing Xylem and Phloem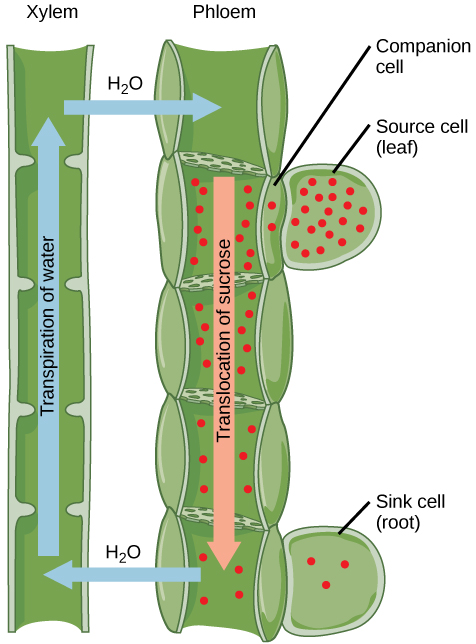
A large oak tree moves hundreds of liters of water from its roots to its highest leaves every single day. No pump pushes this water upward. No heartbeats to drive the flow. Yet water rises continuously against gravity, sometimes to heights of over 100 meters in the tallest trees.
At the same time, sugars produced by photosynthesis in the leaves must reach every living cell in the plant, from the growing tips of roots to the developing seeds. These sugars travel through a completely separate transport system, in any direction the plant requires.
Two systems. Two substances. Two remarkable feats of biological engineering.
Plants have two vascular transport systems running through all their organs.
Both systems run together in vascular bundles throughout the plant.
Transpiration is the loss of water vapor from a plant, primarily through the stomata of leaves.
Transpiration is an unavoidable consequence of having stomata open for gas exchange. When stomata open to allow CO₂ in for photosynthesis, water vapor inevitably diffuses out because the air inside the leaf is saturated with water vapor, while the external air is typically much drier.
The loss of water vapor from leaves by transpiration creates a water potential gradient that pulls water upward through the plant in a continuous stream from roots to leaves. This is called the transpiration stream.
The process works as follows:
This pulling of water upward through xylem by the tension created by transpiration is called the cohesion-tension theory. Water molecules are strongly attracted to each other (cohesion) and to the walls of xylem vessels (adhesion), forming a continuous column that is pulled upward as water is lost at the top.
Xylem vessels are formed from cells that die during development. The cell walls become thickened with lignin and the end walls break down, forming long, hollow tubes with no living contents. Lignin waterproofs the walls and provides structural support.
Water and mineral ions move through xylem by mass flow, the bulk movement of a fluid along a pressure or tension gradient.
Temperature: Higher temperature increases the rate of evaporation of water from mesophyll cell surfaces and increases the kinetic energy of water vapor molecules, speeding diffusion out of the leaf.
Humidity: When the air outside is dry, the water potential gradient between the leaf interior and exterior is steep, and transpiration is rapid. In humid conditions, the gradient is smaller and transpiration slows.
Wind speed: Moving air removes water vapor that accumulates near the leaf surface, maintaining a steep gradient and increasing transpiration rate. Still air allows water vapor to accumulate near the leaf, reducing the gradient.
Light intensity: Light stimulates stomata to open for photosynthesis, increasing the area through which water vapor can escape and therefore increasing transpiration rate.
Leaf surface area: More leaves and larger leaves transpire more water.
Plants in dry habitats have evolved many adaptations to reduce excessive transpiration.
Translocation is the transport of dissolved organic substances through the phloem.
The primary substance transported is sucrose, the form in which carbon is moved through plants. Amino acids, hormones, and other organic molecules are also transported in phloem.
Unlike xylem, which transports water in only one direction (upward), phloem can transport substances in any direction. Sucrose moves from regions where it is produced or released (sources) to regions where it is used or stored (sinks).
Regions that load sucrose into phloem:
Regions that unload sucrose from phloem:
The mechanism of phloem transport is explained by the pressure flow hypothesis.
Phloem consists of two cell types.
Sieve tube elements are living cells with perforated end walls called sieve plates. These allow the flow of phloem contents from cell to cell. Mature sieve tube elements have very few organelles, providing minimal resistance to flow.
Companion cells are closely associated with sieve tube elements and are connected to them by numerous plasmodesmata. They have large numbers of mitochondria and ribosomes, providing the metabolic energy and enzymes needed to actively load and unload sucrose from sieve tubes.
| Feature | Xylem | Phloem |
|---|---|---|
| Contents | Water and mineral ions | Sucrose, amino acids, hormones |
| Direction of flow | Upward only | Any direction |
| Cell type | Dead, hollow vessels | Living sieve tubes |
| Driving force | Transpiration pull (tension) | Pressure flow |
| Energy required | No (passive) | Yes (active loading/unloading) |
| Wall composition | Lignified | Non-lignified |